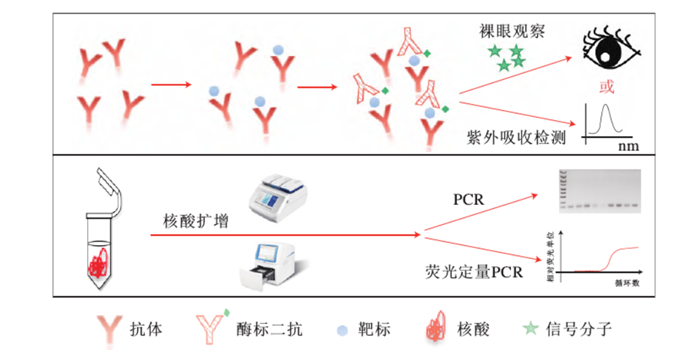
露 心 声 “民以食为天”,食源性致病菌等引起的食物中毒现象,已成为危害公众健康的重要问题。因此,通过食源性致病菌的快速检测提高食品安全保障具有重大意义。今天,小编将带领大家深入了解CRISPR技术在食源性致病菌检测方面的应用。 食品安全问题是关乎人民生命健康和社会和谐的重大战略问题,而食品致病菌是全球范围内导至食品安全问题的主要因素。其中, 以金黄色葡萄球菌、铜绿假单胞菌、单核增生李斯特菌、肠炎沙门氏菌、鲍曼不动杆菌等为代表的食源性致病菌严重威胁着消费者的健康, 由其引起的食源性疾病如腹痛、腹泻以及呕吐等症状近年来呈上升趋势[1],世界卫生组织调查发现,在全球范围内每年约有6亿例食源性疾病发生,其中有42万人因此死亡。因此,尽早的发现食源性致病菌是防止食源性疾病暴发的有效手段。 基于细胞培养的方法尽管一直以来被认为是食源性致病菌检测的金标准,但该方法需要经过增菌、选择性培养、纯化、生化反应鉴定等步骤[2],存在过程繁琐,且检测耗时长等问题,难以满足目前的快速检测需求。而基于抗原抗体特异性结合反应原理的检测方法虽然操作相对简单,但灵敏性不足、容易造成漏检(图1)。因此,开发新型快速高效的食源性致病菌检测技术是亟待解决的重要问题。
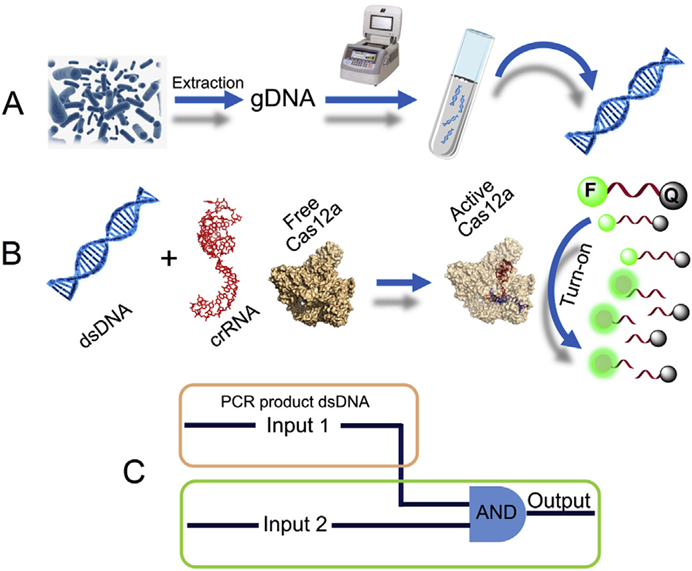
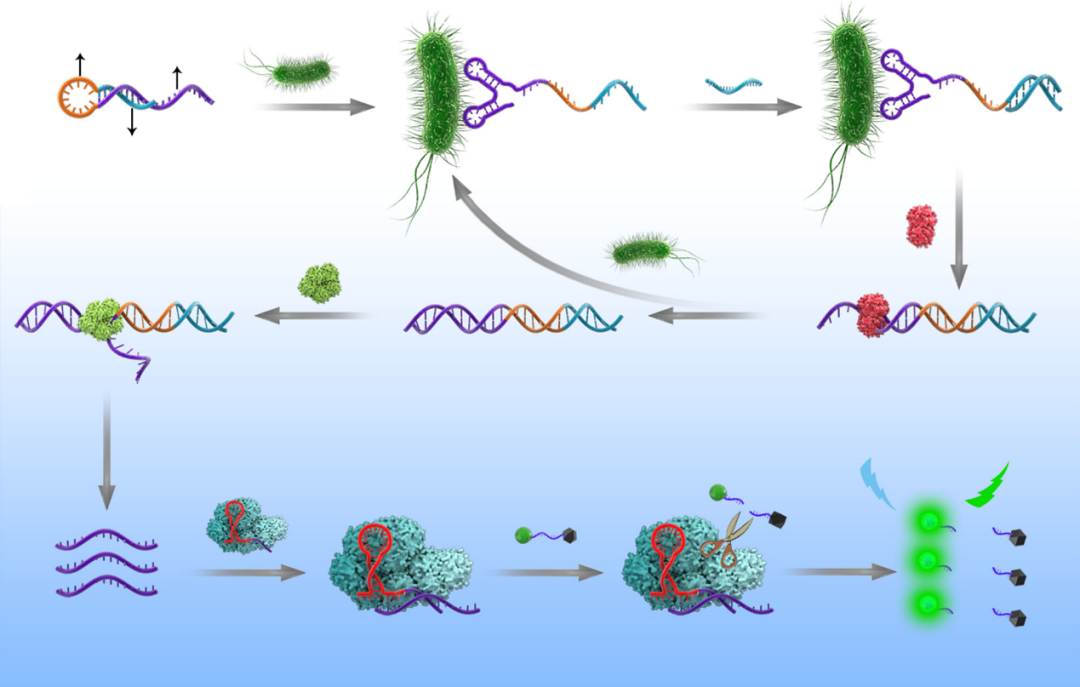
图1. 常规免疫检测及核酸检测原理[3] CRISPR/Cas系统由成簇、规则间隔的短回文重复序列(Clustered Regularly Interspaced Short Palindromic Repeats,CRISPR)和关联蛋白(CRISPR-associated protein,Cas)组成。CRISPR/Cas系统来源于细菌和古细菌的适应性免疫系统[4]。近些年发现的Cas12和Cas13蛋白在CRISPR RNA (crRNA)或向导 RNA (single-guide RNA,sgRNA)引导下,除了具有识别并剪切特异靶标的顺式剪切活性外,还具有可被靶标激活的反式剪切活性。即这些 Cas蛋白与crRNA在特异性识别靶标后,可被激活针对ssDNA或ssRNA的反式剪切活性。基于此原理,可将反式剪切底物设计成合适的报告探针(如荧光-淬灭基团修饰),即可用于靶标核酸检测。 致病菌的CRISPR/Cas核酸传感器检测可分为两大类:①通过增菌后提取基因组,结合核酸扩增和Cas蛋白的剪切体系建立的核酸传感器。②结合适配体实现致病菌向核酸的转化,随后直接或间接与CRISPR/Cas系统结合,建立相应的传感器[5]。在上文中,我们提到了5种常见的食源性致病菌,接下来,编者将以这5种食源性致病菌为例,展开论述现有的CRISPR/Cas检测系统的原理及进展。 金黄色葡萄球菌的检测 Peng[6]等基于PCR和CRISPR/Cas12a建立了PCR-CRISPR/Cas-荧光检测系统,用于金黄色葡萄球菌的检测(图2)。首先通过PCR扩增富集靶标片段,随后将扩增产物与Cas12a/crRNA结合并激活其反式剪切活性,利用该特征剪切荧光探针,以荧光强度的变化指示金黄色葡萄球菌的检测结果,检测线可以达到103 CFU/mL。
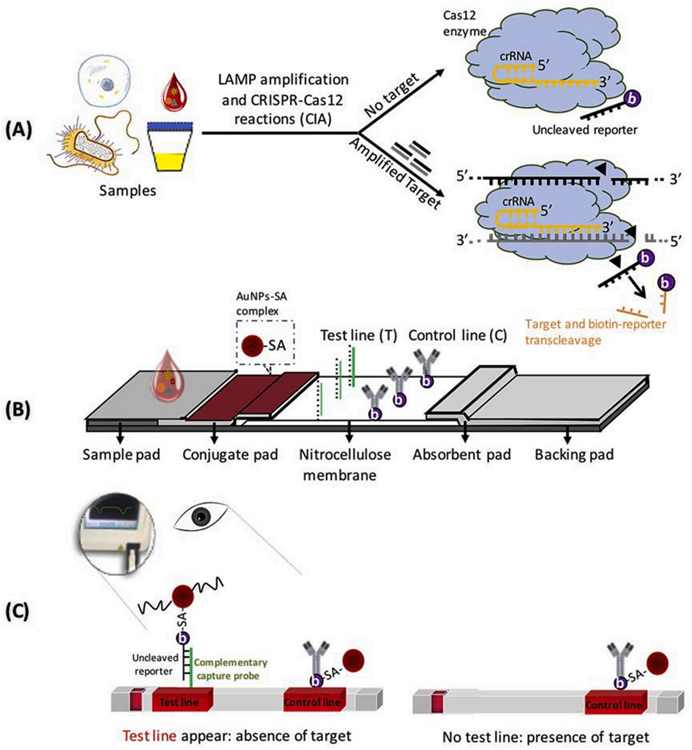
图2. 铜绿假单胞菌的检测 Mukama[7]等基于环介导等温扩增(Loop-mediated isothermal amplification,LAMP)、CRISPR/Cas12a和试纸条技术建立了LAMP-CRISPR/Cas12a-测流层析系统(LFB),用于铜绿假单胞菌的检测(图3)。首先进行LAMP恒温扩增,随后将扩增产物与Cas12a/crRNA结合,启动CRISPR剪切反应。带有biotin的ssDNA被作为Cas12a反式剪切的底物,与报告基因互补的DNA探针序列被固定在LFB的检测线(Test Line)上以捕获报告基因;而生物素偶联的兔多克隆抗IgG抗体被固定在质控线(Control Line)上,以捕获金纳米颗粒-链霉亲和素(AuNP-SA)复合物。当靶标不存在时,Cas12a不会被激活反式剪切活性,带有biotin的ssDNA报告基因维持完整,从而能够被捕获在T线,AuNP-SA偶联物与ssDNA报告基因的biotin结合,并形成AuNP-SA-生物素ssDNA报告复合物,T线显色;反之,靶标存在时,Cas12a会被激活反式剪切活性,剪切带有biotin的ssDNA报告基因,从而T线不显色,AuNP-SA仅在C线上被捕获。通过T线的有无实现对铜绿假单胞菌的定性检测。
图3. LAMP-CRISPR/Cas12a-测流层析系统检测铜绿假单胞菌[7] 核酸试纸及CRISPR试纸定制服务?
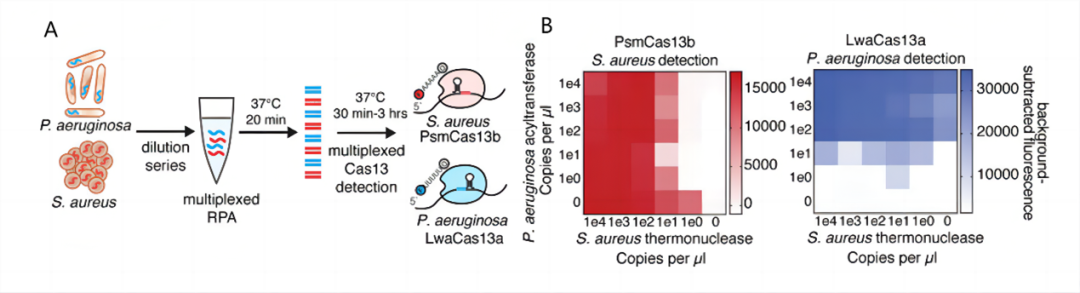
图4. RPA-CRISPR/Cas-双重荧光检测系统对黄色葡萄球菌和铜绿假单胞菌的检测[8] 单核增生李斯特菌的检测 Li[9]等基于重组酶介导的扩增技术(recombinase-aid amplification,RAA)、CRISPR/Cas和电化学技术建立了RAA-CRISPR/Cas-电化学检测系统,用于单核细胞增生李斯特菌的检测。通过进行RAA扩增,然后Cas12a-crRNA双链与靶dsDNA结合,产生一个5′-悬垂的交错剪切。顺式剪切活性的激活进一步激发了Cas12a针对非特异性ssDNA报告基因的反式剪切活性(图5A)。在靶标存在的情况下,Cas12a的反式剪切活性将固定在金电极上的亚甲基蓝标记(MBssDNA)的非特异性ssDNA报告基因从电极表面解离,导至低电化学电流。在没有靶点的情况下,Cas12a/crRNA不会启动非特异性MB-ssDNA报告基因的反式剪切活性,从而产生高电化学电流(图5B),实现了单核增生李斯特菌的检测,最低检测限为26 CFU/mL。
图5. RAA-CRISPR/Cas-电化学传感器检测单核增生李斯特菌[9] 肠炎沙门氏菌的检测 Shen[10]等基于适配体、核酸扩增和CRISPR/Cas13a建立了CRISPR/Cas-荧光传感器,用于肠炎沙门氏菌的免培养检测。Shen等提出了一种APC Cas检测系统,包括基于核酸的变构探针和CRISPR-Cas13a组分,以快速、超灵敏、无DNA提取的策略来直接检测病原体。该系统(如图6)使用了一种特殊的变构探针(AP),是一种单链DNA分子,由三个功能区组成:用于识别目标病原体的适配体功能区(紫色)、引物结合位点功能区(蓝色)和T7启动子功能区(黄色)。此外,在AP的3′端标记一个磷酸基团,以避免自延伸。在没有靶病原体存在的情况下,AP处于具有发夹结构的非活性结合状态,引物结合位点结构域和T7启动子结构域被封闭。在目标病原体存在的情况下,AP的适配体结构域能够特异性识别并与目标病原体结合,导至AP的发夹结构展开,使引物能够退火到引物结合位点。然后,在DNA聚合酶的参与下,AP作为模板产生双链DNA,同时释放病原菌使之能与下一个AP结合,实现病原菌的循环利用。扩增产生的双链DNA在T7 RNA聚合酶的作用下转录产生RNA靶序列,与Cas13a/crRNA结合并激活Cas13的反式剪切活性,剪切荧光报告探针发出荧光信号。因此,APC Cas检测系统通过荧光强度变化实现了肠炎沙门氏菌的快速检测,最低检测限为1 CFU/mL。
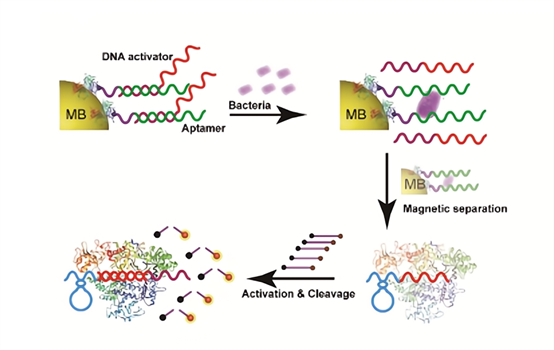
鲍曼不动杆菌的检测 Li [11]等基于适配体和CRISPR/Cas12a建立了CRISPR/Cas-荧光传感器, 用于鲍曼不动杆菌(Acinetobacter baumannii)的免培养免扩增检测。该体系包含针对鲍曼不动杆菌的核酸适配体Aptamer,Aptamer的互补链DNA activator被偶联在磁珠上,当靶标菌体存在时,Aptamer与靶标菌体结合,导至DNA activator被释放,结合Cas12a/crRNA 并激活其反式剪切活性,剪切荧光探针发出信号。该检测平台利用链CRISPR-Cas12a反式剪切的信号放大作用,对鲍曼不动杆菌的最低检测限能达到3 CFU/mL。
图7. CRISPR-Cas12a检测鲍曼不动杆菌[11] 因为CRISPR检测技术具有灵敏、特异、快速、便捷等诸多优势,因此在食源性致病菌检测中具有极大的应用潜力。此外,食品安全检测还包括生物毒素、重金属离子、肉制品成分、非法添加剂等核酸或非核酸靶标,在这些领域也有CRISPR技术的开发和应用,我们也分享过“CRISPR在非核酸靶标检测中的应用”系列导读。 食品安全关乎人人健康,建立更快、更灵敏的食源性致病菌检测方法意义重大!吐露港生物作为CRISPR检测领域的领跑者和专利持有者,愿意全力推动CRISPR诊断产业的快速发展,欢迎联系400-032-6070咨询合作。 |

 /3
/3 

 浙公网安备33010802005999号
浙公网安备33010802005999号