技术|人源抗体开发---B细胞永生化
2020-10-16 09:57|
发布者: 沙糖桔|
查看: 4267|
评论: 0|来源: 体外诊断技术支持 | 作者:ZZY
摘要: 人源单克隆抗体具有免疫原性低、半衰期长等优势,成为了体内应用中不可或缺的生物制剂。人类抗体库为人源单克隆抗体的制备提供了丰富的来源,人B细胞永生化是获得人类抗体库的潜在有效方法,可应用于人源单克隆抗体 ...
| 人源单克隆抗体具有免疫原性低、半衰期长等优势,成为了体内应用中不可或缺的生物制剂。人类抗体库为人源单克隆抗体的制备提供了丰富的来源,人B细胞永生化是获得人类抗体库的潜在有效方法,可应用于人源单克隆抗体的制备。由于各平台均有亟待解决的问题,基于人B细胞永生化的抗体制备尚局限在实验室研究阶段,本文以明确现有的人B细胞永生化抗体制备平台的优劣及其可行性分析,以期为人源单克隆抗体制备技术的进一步发展提供参考。 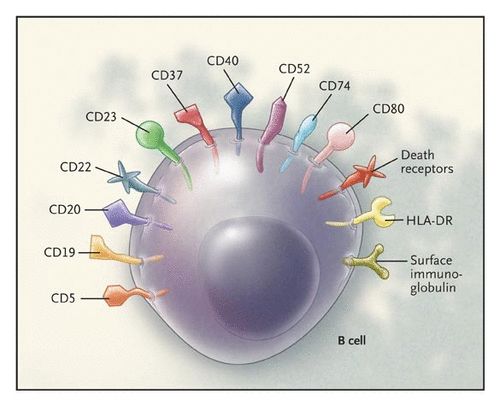
人源单克隆抗体(Human monoclonal antibody,HuMAb) 是指完全由人类基因编码表达的单克隆抗体,已经成为未来抗体发展的主要方向。HuMAb具有更丰富的表位、更低的免疫原性、更长的半衰期、更佳的生物活性,在体内应用中表现良好,如体内诊断、传染病预防、抗体疗法和疾病预后判断等。HuMAb制备过程中还可发掘出高效或/和广泛交叉反应的“超级抗体”,其具有治疗和预防多种病毒感染及其所致疾病的潜力,包括抗原性高度可变的病毒和新兴的或具有大流行潜力的病毒。自SARS-CoV-2引起的新冠肺炎(COVID-19) 疫情爆发以来,全球面临着巨大的公共卫生安全威胁,诊断试剂、抗病毒药物和疫苗的研发迫在眉睫,HuMAb制备技术可以为高效建立新型病毒疫情快速响应平台提供有力的支持。目前,HuMAb的制备主要基于四种技术,分别是转基因动物、抗体库展示、单细胞克隆和B细胞永生化 (表1),各技术可独立进行或者联合使用。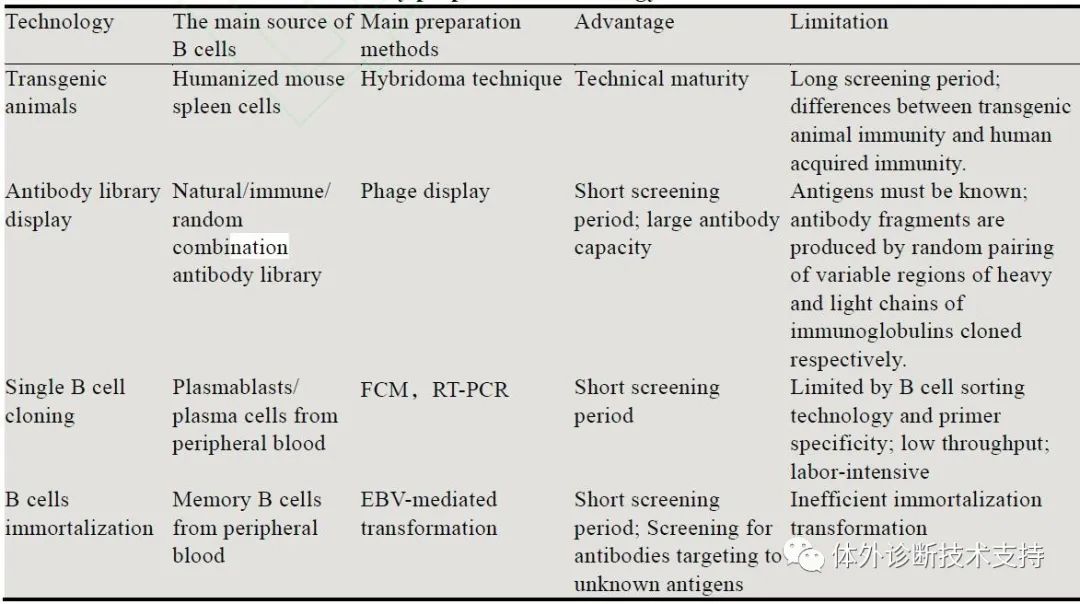
通过转基因动物和抗体库展示技术制备HuMAb已有诸多临床转化的成功实例,但这类抗体不能称之为真正的全人源抗体。小鼠的免疫系统和人类存在很大差异,抗体的亲和成熟、型别转换与人类有差异,无法代表天然人源抗体库。抗体库展示技术利用DNA重组技术将抗体信息展示于特定载体上,由于抗体片段是由分别克隆的免疫球蛋白重链和轻链可变区的随机配对产生的,因此也不能真实反映生理抗体信息。直接从人B细胞中获得目的抗体是一种高效快捷的方法,因为前期无需抗原制备和免疫过程,且天然的抗体库比随机配对的抗体文库具有更高的敏感性和特异性,从受感染者中制备的HuMAb可以提供有关人抗体表位的信息,这些表位对于疫苗的开发以及潜在的治疗应用至关重要。虽然抗体库展示、单细胞克隆和B细胞永生化技术均可利用人B细胞获得HuMAb,但三者差异明显。抗体库展示技术的主要局限如前所述,单B细胞克隆需要先进行繁琐的抗体异源重组表达后才可进行鉴定筛选,B细胞永生化可先进行功能鉴定,直接对抗原特异性阳性的细胞池进行克隆化,与前者相比目的性更强、工作量小、筛选周期更短。综上,与传统方式相比,人B细胞产生的天然抗体经过人类免疫系统选择,是HuMAb的最佳来源,而B 细胞永生化则是获得HuMAb的潜在有效方法。通过B 细胞永生化技术获得的单克隆抗体在生产制备和应用方面具有以下优势: |
声明:
1、凡本网注明“来源:小桔灯网”的所有作品,均为本网合法拥有版权或有权使用的作品,转载需联系授权。
2、凡本网注明“来源:XXX(非小桔灯网)”的作品,均转载自其它媒体,转载目的在于传递更多信息,并不代表本网赞同其观点和对其真实性负责。其版权归原作者所有,如有侵权请联系删除。
3、所有再转载者需自行获得原作者授权并注明来源。

 /3
/3 

 浙公网安备33010802005999号
浙公网安备33010802005999号 









