白细胞介素(Interleukins,ILs)是一大类细胞因子,作为免疫细胞通讯的关键介质,在先天免疫和适应性免疫中调节细胞增殖、分化、运动和存活。 白细胞介素 白细胞介素这一术语在20世纪70年代末首次被引入,用于描述白细胞衍生的细胞因子,其中IL-1被确定为白细胞热原,IL-2被确定为T细胞生长因子。从那时起,超过60种细胞因子被指定为白细胞介素,迄今为止,至少有38种已被正式确认(IL-1至IL-38)。 白细胞介素结构多样,但通常根据受体使用、序列同源性和生物学功能分组为四个家族。
IL-1家族:固有免疫的炎症启动核心 IL-1家族是最早被发现的白细胞介素家族之一,其成员均具有保守的β-三叶折叠结构,受体属于免疫球蛋白(Ig)超家族,信号传导主要依赖NF-κB和MAPK通路激活,核心功能是启动固有免疫应答、调控炎症反应强度,是连接固有免疫与适应性免疫的重要桥梁。与其他白细胞介素家族不同,IL-1家族成员并非仅由免疫细胞分泌,上皮细胞、成纤维细胞等非免疫细胞在应激或损伤状态下也可大量表达,使其成为机体应对外界刺激的“第一道免疫信号枢纽”。
10.1038/s41423-025-01358-8 (一)核心成员组成与特性 目前已发现的IL-1家族成员共11个,包括促炎分子、抗炎分子及功能尚未完全明确的分子,其中核心功能明确的成员主要有:IL-1α、IL-1β、IL-1Ra(IL-1受体拮抗剂)、IL-18、IL-33、IL-36α/β/γ及IL-36Ra等。从分子特性来看,IL-1α和IL-1β是该家族最经典的促炎分子,二者氨基酸序列同源性仅26%,但功能高度重叠,均可结合IL-1受体(IL-1R1)启动炎症信号;不同之处在于,IL-1α可作为膜结合型分子表达于细胞表面,也可通过细胞坏死释放到胞外,而IL-1β则需在炎症小体(如NLRP3炎症小体)的作用下被 caspase-1剪切为成熟体后才具备生物学活性。 IL-1Ra是家族中唯一的抗炎分子,其结构与IL-1α/β高度相似,可竞争性结合IL-1R1,但无法启动信号传导,从而阻断IL-1α/β的促炎作用,是机体维持炎症平衡的关键分子。IL-18和IL-33则属于“警报素”类分子,正常状态下主要储存于细胞内,当细胞受到损伤或感染时释放到胞外,分别调控Th1和Th2型免疫应答,在抗感染和过敏反应中发挥特异性作用。IL-36家族成员(IL-36α/β/γ)主要表达于皮肤、呼吸道黏膜等屏障组织,核心功能是调控局部炎症反应和免疫细胞浸润,而IL-36Ra则作为其特异性拮抗剂,维持局部免疫微环境稳定。 (二)信号传导机制 IL-1家族成员的信号传导均依赖IL-1R家族受体,其中IL-1α/β、IL-36α/β/γ主要结合IL-1R1,IL-18结合IL-1R5(IL-18Rα),IL-33结合IL-1R9(ST2)。以IL-1β为例,其信号传导过程可分为三个核心步骤:第一步,成熟IL-1β与细胞膜表面的IL-1R1结合,诱导IL-1R1构象改变,进而招募共受体IL-1R3(AcP)形成异源二聚体复合物;第二步,复合物通过胞内TIR结构域与衔接蛋白MyD88结合,MyD88进一步招募IRAK家族激酶(IRAK1、IRAK4)形成信号复合物;第三步,IRAK激酶磷酸化激活下游的TRAF6,TRAF6通过泛素化修饰激活TAK1,TAK1进而激活NF-κB和MAPK(ERK、JNK、p38)通路,最终促进促炎因子(如TNF-α、IL-6)、趋化因子(如CXCL8)及黏附分子(如ICAM-1)的基因转录。 IL-1Ra的抗炎机制则是通过竞争性结合IL-1R1实现的,其与IL-1R1的亲和力高于IL-1α/β,结合后可阻止IL-1R1与IL-1R3形成功能性复合物,从而阻断下游信号传导,抑制炎症反应的过度激活。IL-18和IL-33的信号传导过程与IL-1β类似,均依赖MyD88-IRAK-TRAF6信号轴,但下游激活的转录因子存在差异:IL-18主要激活STAT1和NF-κB,促进IFN-γ分泌;IL-33则主要激活STAT6和NF-κB,促进IL-4、IL-5、IL-13等Th2型细胞因子分泌。 (三)核心生物学功能 1. 启动固有免疫应答:IL-1α/β是机体炎症反应的“首发因子”,当机体受到细菌、病毒感染或组织损伤时,巨噬细胞、中性粒细胞等固有免疫细胞首先分泌IL-1α/β,通过诱导内皮细胞表达黏附分子(ICAM-1、VCAM-1),促进中性粒细胞、单核细胞向炎症部位浸润,同时激活巨噬细胞分泌更多促炎因子,形成炎症放大效应,快速清除病原体或修复损伤组织。在感染早期,IL-1α/β还可诱导树突状细胞(DC)成熟,增强其抗原提呈能力,为适应性免疫应答的启动奠定基础。 2. 调控适应性免疫分化:IL-18与IL-12协同作用,可显著促进Th0细胞向Th1细胞分化,并增强Th1细胞和NK细胞分泌IFN-γ,强化机体抗胞内病原体(如结核分枝杆菌、病毒)和抗肿瘤免疫应答;此外,IL-18还可促进CTL细胞的活化和增殖,提升其杀伤靶细胞的能力。IL-33则主要诱导Th0细胞向Th2细胞分化,促进B细胞分泌IgE,同时激活肥大细胞和嗜酸性粒细胞,参与过敏反应和抗寄生虫感染免疫;在组织损伤修复过程中,IL-33还可促进成纤维细胞增殖和胶原合成,加速组织愈合。 3. 维持炎症平衡:IL-1Ra与IL-1α/β的动态平衡是维持机体炎症稳态的关键。正常生理状态下,IL-1Ra的表达水平较低,当炎症反应启动后,IL-1Ra可通过自分泌或旁分泌方式大量表达,与IL-1α/β竞争结合IL-1R1,避免炎症反应过度激活对正常组织造成损伤。此外,IL-36Ra通过阻断IL-36家族成员的信号传导,可有效抑制皮肤、呼吸道等黏膜组织的过度炎症反应,预防自身免疫性疾病的发生。 共用γ链家族:适应性免疫的发育与活化枢纽 共用γ链家族(Common γ Chain Family,γc家族)属于Ⅰ型细胞因子受体超家族的核心亚类,其最显著的特征是所有成员的受体均包含共同的信号亚基——γ链(γc,CD132)。γc是细胞因子信号传导的关键分子,通过与不同的配体结合亚基形成异源二聚体或三聚体受体,介导JAK3-STAT信号通路激活,核心功能是调控T细胞、B细胞、NK细胞等适应性免疫细胞的发育、存活、增殖及活化,是维持机体适应性免疫功能完整的核心家族。 (一)核心成员组成与受体特征
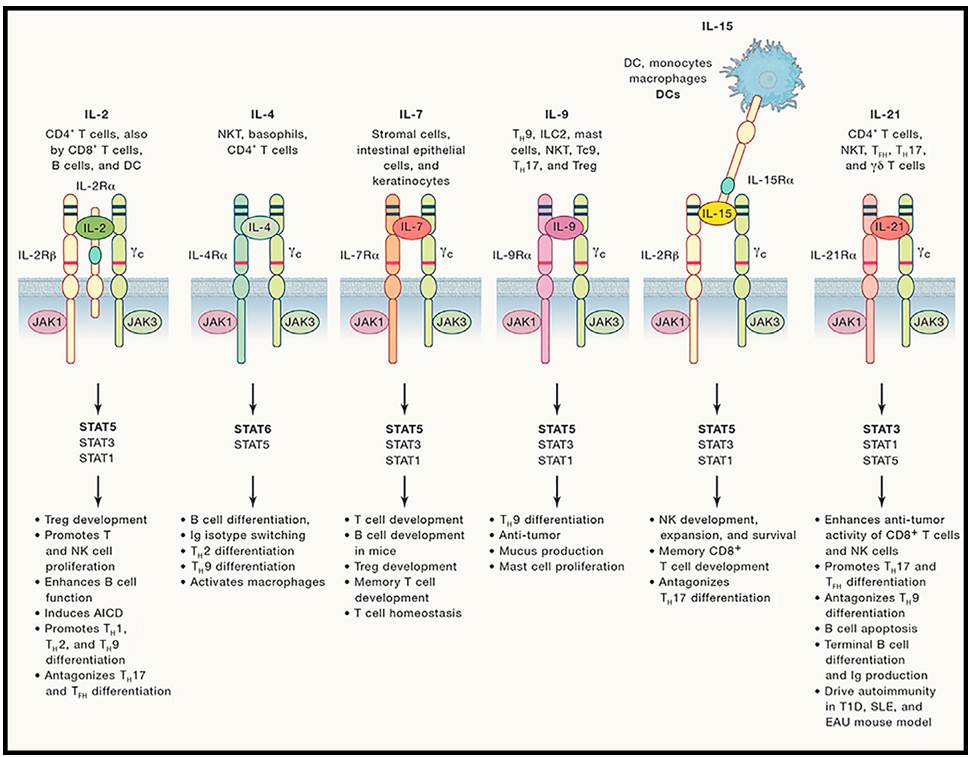
10.1016/j.immuni.2019.03.028. 共用γ链家族的核心成员包括IL-2、IL-4、IL-7、IL-9、IL-15及IL-21,这些成员的氨基酸序列同源性较低,但受体均以γc作为共同信号亚基。根据受体组成的差异,可将家族成员分为两类:一类是受体由配体结合亚基与γc组成的异源二聚体,包括IL-4(受体为IL-4Rα+γc)、IL-7(IL-7Rα+γc)、IL-9(IL-9Rα+γc);另一类是受体由配体结合亚基、β链(βc,CD131)与γc组成的异源三聚体,包括IL-2(IL-2Rα+IL-2Rβ+γc)、IL-15(IL-15Rα+IL-2Rβ+γc)。IL-21的受体则由IL-21Rα与γc组成异源二聚体,其信号传导机制与二聚体受体家族成员类似。 值得注意的是,IL-2Rα(CD25)仅负责结合IL-2,不参与信号传导,而IL-2Rβ(CD122)与γc则是信号传导的核心亚基;IL-15Rα则可作为“载体”分子,与IL-15结合后通过转呈作用(Trans-presentation)将IL-15传递给表达IL-2Rβ+γc的细胞,这种转呈机制可显著增强IL-15的信号传导效率和特异性。此外,家族成员的表达细胞具有明显的特异性:IL-2主要由活化的CD4+ T细胞和CD8+ T细胞分泌;IL-4主要由Th2细胞、肥大细胞和嗜碱性粒细胞分泌;IL-7主要由胸腺基质细胞、骨髓基质细胞和成纤维细胞分泌;IL-9由Th9细胞分泌;IL-15由巨噬细胞、树突状细胞和上皮细胞分泌;IL-21则主要由Tfh细胞和NK细胞分泌。 (二)信号传导机制 共用γ链家族成员的信号传导均依赖JAK3-STAT通路,其核心机制具有高度一致性,具体过程以IL-2为例:第一步,IL-2与细胞膜表面的IL-2Rα结合,诱导IL-2Rα与IL-2Rβ+γc形成三聚体受体复合物;第二步,受体复合物的形成可促使γc胞内结构域结合的JAK3与IL-2Rβ结合的JAK1发生相互磷酸化激活;第三步,激活的JAK1和JAK3可磷酸化受体胞内的酪氨酸残基,形成STAT分子的结合位点;第四步,STAT分子(如STAT5、STAT3、STAT6)结合到磷酸化的酪氨酸残基上,被JAK激酶磷酸化激活,激活的STAT分子形成同源二聚体或异源二聚体,进入细胞核内;第五步,STAT二聚体结合到靶基因的启动子区域,调控基因转录,如STAT5可促进Bcl-2、Cyclin D2等抗凋亡和促增殖基因的表达,STAT6则可促进GATA3等Th2细胞分化相关基因的表达。 不同家族成员激活的STAT分子存在差异,这也是其功能特异性的核心原因:IL-2、IL-7、IL-15主要激活STAT5,调控免疫细胞的存活和增殖;IL-4主要激活STAT6,调控Th2细胞分化和IgE合成;IL-21主要激活STAT3,调控Tfh细胞分化和B细胞活化。此外,除了JAK3-STAT通路,家族成员还可激活PI3K-AKT通路,进一步增强免疫细胞的增殖和存活能力。需要注意的是,γc是JAK3的特异性结合伙伴,若γc或JAK3发生突变,将导至整个家族的信号传导受阻,引发严重的联合免疫缺陷病(SCID),这也充分证明了该家族在免疫发育中的核心作用。 (三)核心生物学功能 1. 调控T细胞发育与稳态:IL-7是T细胞发育过程中的关键因子,在胸腺中,IL-7可促进早期胸腺细胞(DN细胞)的增殖和分化,维持胸腺微环境的稳态;对于外周初始T细胞,IL-7可通过激活STAT5通路,促进Bcl-2的表达,抑制细胞凋亡,维持初始T细胞的存活。IL-2则是活化T细胞增殖和分化的核心因子,活化的T细胞通过自分泌IL-2,促进自身增殖分化为效应T细胞(如Th1、Th2、Th17细胞)和记忆T细胞;同时,IL-2还可促进调节性T细胞(Treg)的增殖和功能维持,通过抑制过度的免疫应答,维持免疫耐受。IL-15则可促进记忆CD8+ T细胞的形成和存活,在抗病毒和抗肿瘤免疫的长期保护中发挥关键作用。 2. 调控B细胞发育与活化:IL-4是B细胞活化和分化的重要调控因子,可诱导B细胞向IgE分泌型浆细胞分化,同时促进B细胞表达MHCⅡ类分子和共刺激分子(CD80、CD86),增强其抗原提呈能力。IL-21则主要调控Tfh细胞与B细胞的相互作用,促进B细胞在生发中心的增殖、分化,形成高亲和力浆细胞和记忆B细胞,参与抗体的 affinity maturation 过程。此外,IL-7也可促进骨髓中B细胞前体的发育,维持B细胞池的稳态。 3. 调控NK细胞功能:IL-15是NK细胞发育、增殖和活化的关键因子,在骨髓中,IL-15可促进NK细胞前体的分化成熟;在外周,IL-15可激活NK细胞,增强其对靶细胞的杀伤能力,同时促进NK细胞分泌IFN-γ等细胞因子,参与固有免疫应答。IL-2也可激活NK细胞,增强其杀伤活性,是临床中用于激活NK细胞抗肿瘤的重要因子。IL-21则可调控NK细胞的增殖和功能,增强其在肿瘤微环境中的存活能力和杀伤效率。 IL-6家族:炎症平衡与多系统调控的核心因子 IL-6家族同样属于Ⅰ型细胞因子受体超家族,其核心特征是所有成员的受体均包含共同的信号亚基——糖蛋白130(gp130,CD130)。gp130是一种广泛表达于多种细胞表面的跨膜蛋白,通过与不同的配体结合亚基形成二聚体或四聚体受体,介导JAK-STAT3、MAPK等多条信号通路的激活。IL-6家族成员兼具促炎、抗炎、免疫调控、造血支持及组织修复等多重功能,调控范围覆盖免疫、造血、神经、代谢等多个系统,是连接炎症反应与多器官功能稳态的关键家族。 (一)核心成员组成与受体特征
10.1136/jitc-2023-007530 IL-6家族的核心成员包括IL-6、IL-11、IL-27、IL-31、IL-35、睫状神经营养因子(CNTF)、心肌营养素-1(CT-1)等,其中IL-6、IL-11、IL-27、IL-31、IL-35是免疫调控的核心成员。根据受体组成的差异,可将家族成员分为两类:一类是受体由配体结合亚基与gp130组成的异源二聚体,包括IL-11(受体为IL-11Rα+gp130)、IL-31(IL-31Rα+gp130);另一类是受体由两个不同的配体结合亚基与两个gp130组成的异源四聚体,包括IL-6(IL-6Rα+gp130)、IL-27(IL-27Rα+gp130)、IL-35(IL-35Rα+gp130)。 IL-6是家族中最经典的成员,其受体IL-6Rα有两种形式:膜结合型IL-6Rα(mIL-6Rα)和可溶性IL-6Rα(sIL-6Rα)。mIL-6Rα主要表达于肝细胞、巨噬细胞、T细胞等细胞表面,而sIL-6Rα可通过蛋白酶剪切mIL-6Rα产生或直接由细胞分泌,sIL-6Rα可与IL-6结合形成复合物,再与表达gp130的细胞结合启动信号传导,这种“跨信号传导”机制扩大了IL-6的作用范围,使其可调控不表达mIL-6Rα的细胞(如内皮细胞、平滑肌细胞)。IL-27和IL-35则是由两个亚基组成的异源二聚体分子:IL-27由EBI3和p28亚基组成,IL-35由EBI3和p35亚基组成,这种独特的结构使其兼具促炎和抗炎的双重调控功能。 (二)信号传导机制
10.1111/febs.17224 IL-6家族成员的信号传导以JAK-STAT3通路为主,同时可激活MAPK和PI3K-AKT通路,其核心机制以IL-6为例:第一步,IL-6与mIL-6Rα或sIL-6Rα结合形成复合物;第二步,复合物与细胞膜表面的gp130结合,诱导两个gp130分子形成同源二聚体;第三步,gp130胞内结构域结合的JAK家族激酶(JAK1、JAK2、Tyk2)发生相互磷酸化激活;第四步,激活的JAK激酶磷酸化gp130胞内的酪氨酸残基,形成STAT3的结合位点;第五步,STAT3结合到磷酸化的酪氨酸残基上,被JAK激酶磷酸化激活,激活的STAT3形成同源二聚体进入细胞核内;第六步,STAT3结合到靶基因的启动子区域(如急性期反应基因、促炎因子基因、抗凋亡基因),调控基因转录。 不同家族成员激活的信号通路和转录因子存在差异:IL-6主要激活STAT3和MAPK通路,调控急性期反应和炎症反应;IL-27可激活STAT1和STAT3通路,其中STAT1介导其抗炎功能,STAT3介导其促炎功能;IL-35则主要激活STAT1和STAT4通路,通过抑制促炎因子的表达发挥抗炎作用;IL-31主要激活STAT3和STAT5通路,参与瘙痒和炎症反应的调控。此外,家族成员的信号传导还受到负向调控分子的调控,如SOCS3(细胞因子信号抑制因子3)可通过与磷酸化的gp130结合,抑制JAK激酶的活性,从而负向调控信号传导,维持信号稳态。 (三)核心生物学功能 1. 调控急性期炎症反应:IL-6是机体最重要的急性期反应因子之一,当机体受到细菌感染、组织损伤或应激刺激时,巨噬细胞、树突状细胞等固有免疫细胞快速分泌IL-6,通过激活肝细胞表面的IL-6Rα+gp130受体,诱导肝脏合成C反应蛋白(CRP)、纤维蛋白原、血清淀粉样蛋白A(SAA)等急性期蛋白。这些急性期蛋白可通过激活补体系统、促进吞噬细胞的吞噬功能,增强机体对病原体的清除能力;同时,IL-6还可通过跨信号传导机制,诱导内皮细胞表达黏附分子,促进免疫细胞向炎症部位浸润,放大炎症反应。 2. 免疫分化的双向调控:IL-6家族成员在适应性免疫细胞的分化中发挥关键的双向调控作用。IL-6与TGF-β协同作用,可促进Th0细胞向Th17细胞分化,Th17细胞分泌的IL-17等促炎因子参与自身免疫病的发病;同时,IL-6还可抑制Treg细胞的分化和功能,进一步加重免疫紊乱。IL-27则具有双重调控功能:一方面,IL-27可促进初始T细胞向Th1细胞分化,增强抗胞内病原体免疫;另一方面,IL-27可抑制Th17细胞和Th2细胞的分化,促进调节性T细胞的生成,维持免疫耐受。IL-35则是经典的抗炎分子,主要由Treg细胞分泌,通过抑制Th1、Th17细胞的增殖和功能,以及抑制巨噬细胞、树突状细胞的活化,发挥免疫抑制作用,在自身免疫病和肿瘤免疫逃逸中发挥重要作用。 3. 造血支持与组织修复:IL-11是重要的造血调控因子,可促进骨髓造血干细胞的增殖和分化,尤其是促进巨核细胞的成熟和血小板的生成,已被用于治疗化疗后血小板减少症。IL-6也可促进造血干细胞的增殖和分化,增强骨髓的造血功能。在组织修复过程中,IL-6可促进成纤维细胞的增殖和胶原合成,加速伤口愈合;CT-1等家族成员则可促进心肌细胞的存活和修复,在心血管疾病的病理生理过程中发挥作用。 IL-17家族:黏膜免疫与炎症损伤的关键调控因子 IL-17家族是一类结构独特的促炎细胞因子家族,其成员均以同源二聚体或异源二聚体的形式存在,受体属于IL-17R家族。IL-17家族的核心功能是介导黏膜免疫防御和炎症组织损伤,通过诱导上皮细胞、成纤维细胞等分泌趋化因子和抗菌肽,招募中性粒细胞到炎症部位,在细菌、真菌等病原体的黏膜感染防御中发挥关键作用;但过度激活则会导至组织炎症损伤,与多种自身免疫性疾病的发病机制密切相关。此外,IL-17家族成员在肿瘤免疫中发挥双重作用,其功能取决于肿瘤类型和肿瘤微环境。
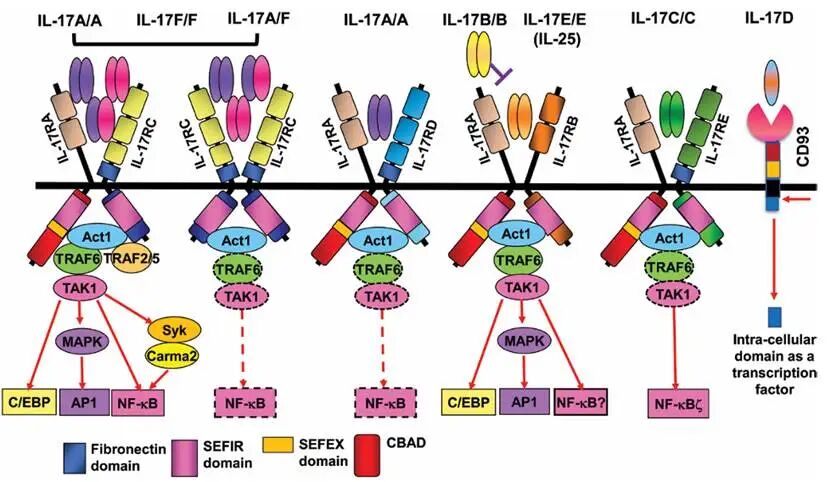
10.1093/intimm/dxab075. (一)核心成员组成与受体特征 目前已发现的IL-17家族成员共6个,分别为IL-17A、IL-17B、IL-17C、IL-17D、IL-17E(即IL-25)和IL-17F。其中,IL-17A和IL-17F的氨基酸序列同源性高达50%,均以同源二聚体(IL-17A/A、IL-17F/F)或异源二聚体(IL-17A/F)的形式存在,是家族中功能最相似的两个成员。IL-17E(IL-25)的结构和功能与其他成员差异较大,更倾向于调控Th2型免疫应答,参与过敏反应和寄生虫感染。 IL-17家族的受体属于IL-17R家族,包括IL-17RA、IL-17RB、IL-17RC、IL-17RD、IL-17RE五个成员,均为Ⅰ型跨膜蛋白,胞内含有保守的SEFIR结构域。不同IL-17家族成员与IL-17R家族成员的结合具有特异性:IL-17A、IL-17F及IL-17A/F主要结合IL-17RA/RC异源二聚体受体;IL-17B结合IL-17RB;IL-17C结合IL-17RA/RE;IL-17D的受体尚未完全明确,可能结合IL-17RA/RD;IL-17E(IL-25)结合IL-17RB/RE。IL-17R家族受体的表达具有明显的组织特异性,IL-17RA广泛表达于多种细胞表面,而IL-17RB、RC、RE则主要表达于上皮细胞、黏膜组织细胞及免疫细胞表面,这种特异性表达决定了IL-17家族成员的功能特异性。 从表达细胞来看,IL-17A和IL-17F主要由Th17细胞分泌,此外,γδT细胞、NK细胞、固有淋巴细胞(ILC3)等也可分泌;IL-17B主要由成纤维细胞、上皮细胞分泌;IL-17C主要由上皮细胞分泌,是一种“上皮源性”细胞因子;IL-17D主要由T细胞、NK细胞分泌;IL-17E(IL-25)主要由上皮细胞、肥大细胞、嗜酸性粒细胞分泌。 (二)信号传导机制 IL-17家族成员的信号传导主要依赖NF-κB和MAPK通路,其核心机制以IL-17A为例:第一步,IL-17A同源二聚体与细胞膜表面的IL-17RA/RC异源二聚体受体结合;第二步,受体结合后,胞内的SEFIR结构域招募衔接蛋白ACT1(也称为CIKS);第三步,ACT1通过其U-box结构域发挥E3泛素连接酶活性,催化自身及下游分子的泛素化修饰;第四步,泛素化修饰的ACT1激活下游的TAK1和IKK复合物;第五步,TAK1激活MAPK(ERK、JNK、p38)通路,IKK复合物激活NF-κB通路;第六步,两条通路共同调控促炎因子(如IL-6、TNF-α)、趋化因子(如CXCL8、CXCL1)及抗菌肽(如β-防御素、S100蛋白)的基因转录。 IL-17E(IL-25)的信号传导机制与IL-17A类似,但下游激活的转录因子存在差异,其主要激活NF-κB和STAT6通路,促进IL-4、IL-5、IL-13等Th2型细胞因子的分泌,调控Th2型免疫应答。此外,IL-17家族成员的信号传导还可通过与其他细胞因子(如TNF-α、IL-6)的协同作用,放大炎症反应,例如,IL-17A与TNF-α协同作用可显著增强促炎因子和趋化因子的分泌。 (三)核心生物学功能 1. 黏膜免疫防御:IL-17家族成员是黏膜免疫防御的关键因子,尤其在肠道、呼吸道、皮肤等黏膜屏障组织的抗感染中发挥重要作用。IL-17A和IL-17F可诱导肠道、呼吸道上皮细胞分泌CXCL8、CXCL1等趋化因子,招募中性粒细胞到感染部位,清除细菌、真菌等病原体;同时,可诱导上皮细胞分泌β-防御素、S100A8/A9等抗菌肽,直接杀灭病原体。IL-17C作为上皮源性细胞因子,可通过自分泌或旁分泌方式激活上皮细胞,增强黏膜屏障功能,在肠道炎症和感染中发挥保护作用。IL-17E(IL-25)则主要参与抗寄生虫感染免疫,通过诱导Th2型免疫应答,促进嗜酸性粒细胞活化和IgE分泌,清除寄生虫。 2. 介导炎症损伤:IL-17家族成员的过度激活是导至多种自身免疫性疾病炎症损伤的关键原因。在银屑病患者的皮损组织中,IL-17A的表达水平显著升高,通过诱导角质形成细胞分泌促炎因子和抗菌肽,促进中性粒细胞浸润,形成典型的银屑病皮损(红斑、鳞屑);在强直性脊柱炎患者的滑膜组织和关节液中,IL-17A和IL-17F的表达异常升高,通过促进滑膜炎症和骨破坏,加重关节损伤。此外,IL-17家族成员还与类风湿关节炎、多发性硬化、炎症性肠病等疾病的炎症进展密切相关。 3. 肿瘤免疫的双重作用:IL-17家族成员在肿瘤免疫中发挥复杂的双重作用。一方面,IL-17可通过诱导炎症微环境,促进肿瘤血管生成和肿瘤细胞的侵袭转移,同时抑制抗肿瘤免疫细胞(如CD8+ T细胞)的功能,促进肿瘤进展;例如,在结直肠癌、肺癌等肿瘤中,IL-17A的高表达与肿瘤分期进展和不良预后相关。另一方面,IL-17可激活固有免疫细胞(如中性粒细胞、NK细胞),增强其抗肿瘤活性,同时促进CD8+ T细胞的浸润和活化,抑制肿瘤生长;例如,在黑色素瘤、肝癌等肿瘤中,IL-17的表达与患者的良好预后相关。这种双重作用的具体机制尚未完全明确,可能与肿瘤类型、肿瘤微环境中免疫细胞的组成及IL-17家族成员的亚型有关。 参考资料 Cecilia Garlanda et al, IL-1 family cytokines in inflammation and immunity, Cellular & Molecular Immunology (2025) 22:1345–1362; https://doi.org/10.1038/s41423-025-01358-8 The γc Family of Cytokines: Basic Biology to Therapeutic Ramifications,Immunity. 2019 Apr 16;50(4):832-850. doi: 10.1016/j.immuni.2019.03.028. Yadav MK, Singh SP and Egwuagu CE (2025) IL-6/IL-12 superfamily of cytokines and regulatory lymphocytes play critical roles in the etiology and suppression of CNS autoimmune diseases. Front. Immunol. 16:1514080. doi: 10.3389/fimmu.2025.1514080 Ritzmann, F.; Lunding, L.P.; Bals, R.; Wegmann, M.; Beisswenger, C. IL-17 Cytokines and Chronic Lung Diseases. Cells 2022, 11, 2132. https:// doi.org/10.3390/cells11142132 Alexandra A Wolfarth et al, Advancements of Common Gamma-Chain Family Cytokines in Cancer Immunotherapy, Immune Netw. 2022 Feb 22;22(1):e5. doi: 10.4110/in.2022.22.e5 |

 /3
/3 

 浙公网安备33010802005999号
浙公网安备33010802005999号