摘要 尽管近年来微生物基因组数据激增,但通过基于培养的实验对于证实细胞生物学、生态作用和微生物进化的推论仍然十分重要。目前绝大多数古菌和细菌仍难以培养且对其特性了解不够充分,因此研究者们对高效的培养学方法愈加重视,这也加快了许多方法学和技术的发展。本综述讨论了可能阻碍新型微生物的分离和培养的常见障碍,并介绍了具有针对性或高通量培养的新兴技术。此外,还重点介绍了成功培养新型古细菌和细菌的最新实例,并提出了未来可尝试培养的关键微生物。 编译:鞠志成 英文标题: 中文标题: 期刊: Nature Reviews Microbiology,2020 第一作者: 通讯作者: Thijs J. G. Ettema 作者单位: 前言 基因组测序技术、复杂的宏基因组学及系统发育学的进步极大地改变了我们对微生物生命多样性的理解,其中就包括对“生命之树”形状的描绘。但目前对于古细菌和细菌的了解,要么来自少数经过精心研究的培养谱系,要么来自于未培养谱系的重建基因组。因此对一些新颖谱系的代表微生物进行纯培养十分必要,并至少具有以下三点意义:
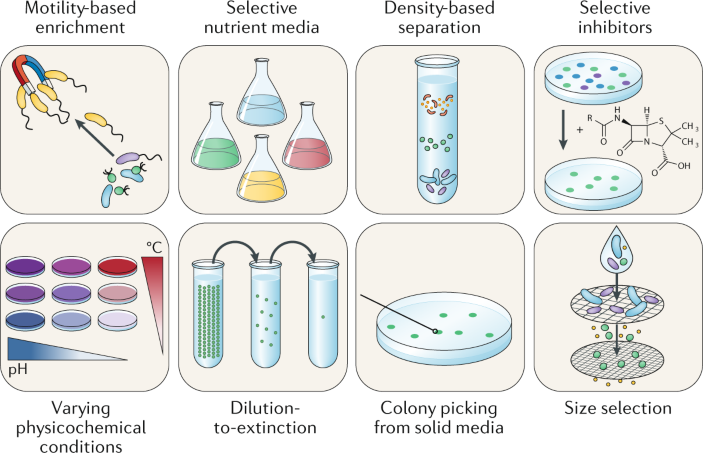
正文 01 经典培养方法 微生物培养起源于19世纪中叶,许多现代的富集分离方法都借鉴了一个多世纪以前引入的某些原理,其中许多依赖于直接观察培养物的生理行为以及所含微生物的表型和基因型特征(Fig 1)。灵活运用这些方法不仅需要消耗大量的时间和耐心,还很大程度依赖于研究人员的经验判断。
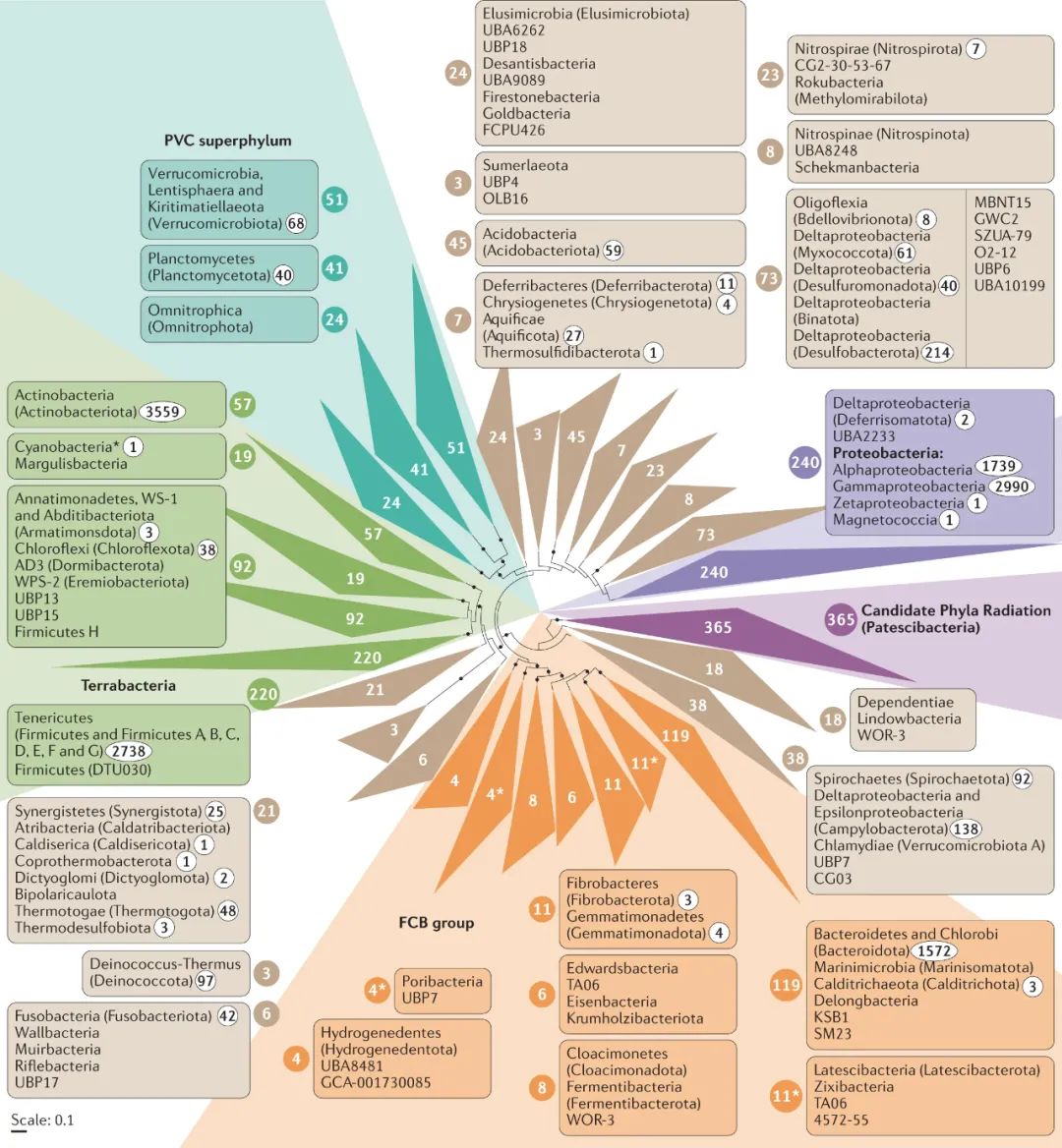
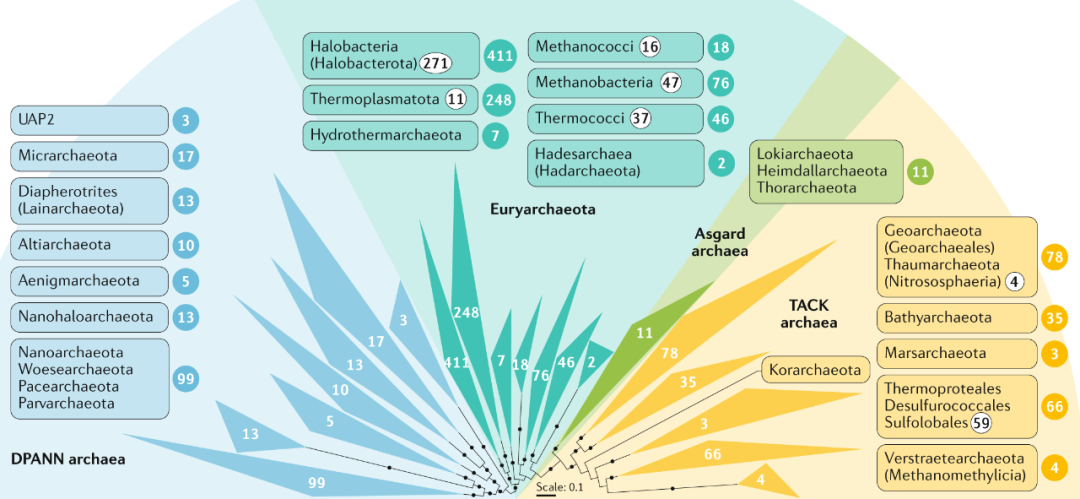
Figure 1. 常见的传统微生物纯培养方法 02 解释“The uncultured majority” “生命之树”是生物学中最重要的概念之一,在过去的几十年中已得到了广泛的扩展,其中包括几个具有较高分类学等级的古细菌和细菌。通常把生命之树中的原核生物分为两个域,即古细菌域和细菌域。据估计它们共包含数百至数千个门,随着基因组数据的积累,这一数字仍在不断增加(尽管估算方法可能不同)。根据16S rRNA基因序列数据,已计算出古菌和细菌的总数约为400,000个,包括约60,000个属,尽管其实际数量可能超出估算值几个数量级。然而只有约14000古细菌和细菌物种(分布在3500属和38个门)已被培育和有效描述。在这些物种中,约97%仅属于四个细菌门(拟杆菌门,变形菌门,厚壁菌门和放线菌门)(Fig 2)。相反,所有其他细菌门以及整个古菌都由相对较少的纯培养物种所代表(Fig 2、3)。
Figure 2. 已培养细菌主要是拟杆菌门、变形杆菌门、厚壁菌门和放线菌门 从每个物种总共15种核糖体蛋白的至少5种的串联比对中,推断出细菌的系统发育树,根据基因组分类数据库获得的1,541个细菌基因组进行编码。彩色圆圈中的白色数字是每个折叠的进化枝中各个分类单元的数目,也用于将相应的分类单元名称连接到进化枝。基于BacDive数据库中存在的分配给每个进化枝的物种类型菌株的数量,分类单元名称旁边的白圈黑字表示针对这些分类单元描述的培养分离株的物种数(数据收集截止2020.4.6)。
Figure 3. 古细菌的多样性主要由未培养的群体主导 从每个物种总共 |

 /3
/3 

 浙公网安备33010802005999号
浙公网安备33010802005999号