登陆有奖并可浏览互动!
您需要 登录 才可以下载或查看,没有账号?立即注册


×
细胞炎性坏死 (焦亡) 常伴随着线粒体的损伤,这一切的背后到底是道德的沦丧还是……咳咳咳,这究竟是巧合还是早有预谋?让我们随着小 M 来探索一下其中的奥秘吧~
 细胞焦亡又称细胞炎症性坏死,一种程序性细胞死亡方式,由炎症小体的活化及炎症性半胱天冬酶的激活引起,是机体免疫系统对抗外源病原菌入侵的主要手段之一[1][2][3][4]。往期研究发现 GSDMD/E 介导的细胞焦亡早期伴随线粒体损伤发生,但其潜在机制和功能尚不清楚[5]。 细胞焦亡又称细胞炎症性坏死,一种程序性细胞死亡方式,由炎症小体的活化及炎症性半胱天冬酶的激活引起,是机体免疫系统对抗外源病原菌入侵的主要手段之一[1][2][3][4]。往期研究发现 GSDMD/E 介导的细胞焦亡早期伴随线粒体损伤发生,但其潜在机制和功能尚不清楚[5]。
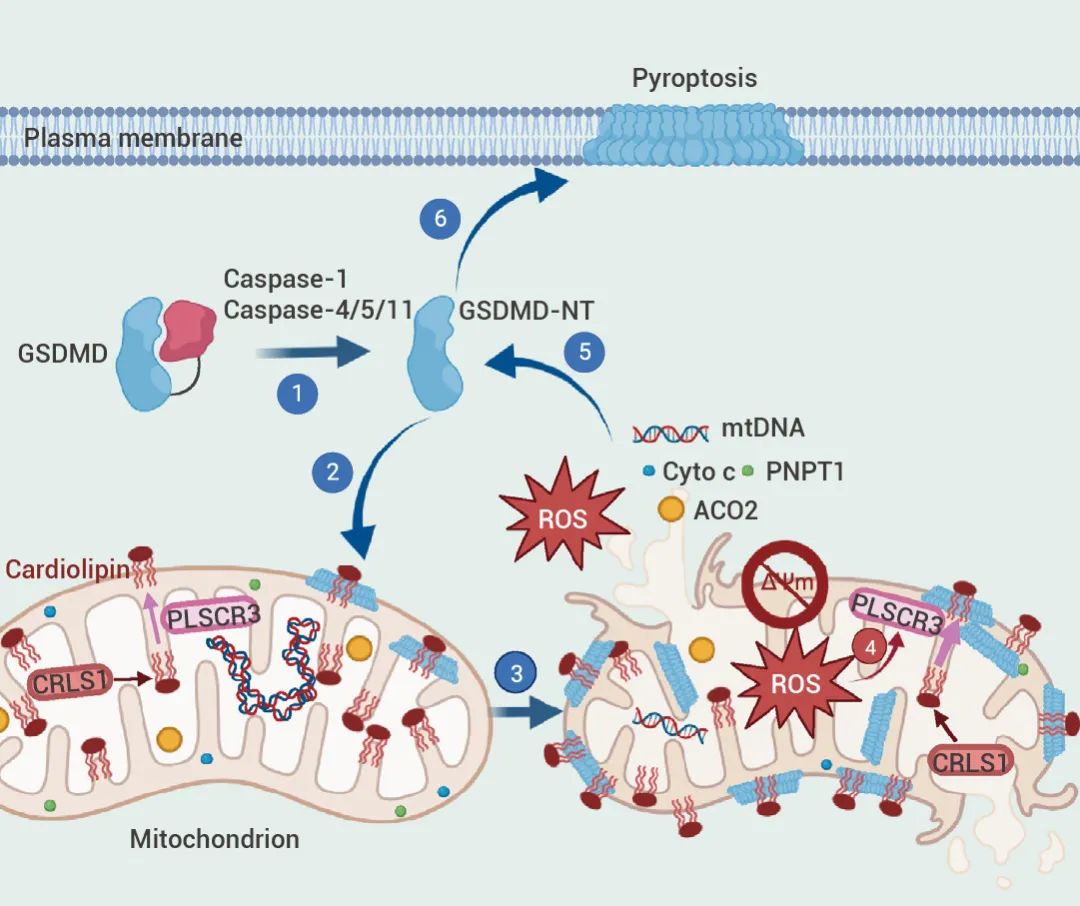
近期,Miao 等人发表在 Immunity 的研究首次报道了 GSDMD 介导线粒体损伤的分子机制!对细胞死亡和炎症来说,线粒体损伤是一条"不归路":激活的 Gasdermins 结合到线粒体的心磷脂,形成线粒体孔道,这将破坏两个线粒体膜,导至活性氧的产生,氧化磷酸化的丧失,细胞毒性介质的释放,以及线粒体自噬[1]。GSDMD 介导线粒体损伤的分子机制[1]。 科学不废话!各位看官,来一来,瞅一瞅,走过路过不要错过哈哈哈,让我们一起浏览一下吧~~

▐ 线粒体受损:评估线粒体敏感性和膜电位
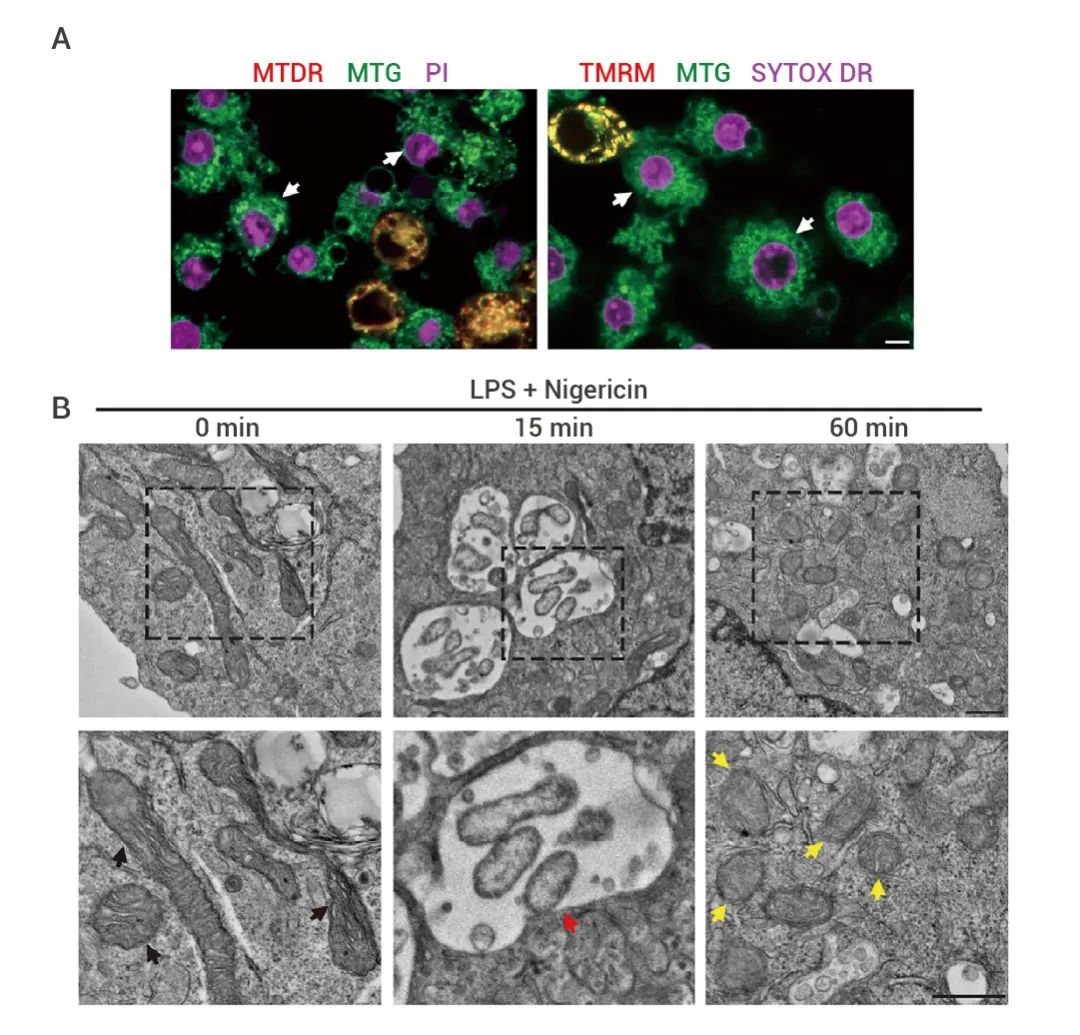
研究人员使用 LPS + Nigericin 联合处理 THP-1 巨噬细胞,结合染料 MTDR、MTG 以及 TMRM 进行染色,评估线粒体膜的电位和完整性 (图 1A-B)。与邻近的 SYTOX 或 PI 阴性活细胞相比,SYTOX 或 PI 阳性的焦亡细胞的 MTDR 和 TMRM 荧光明显减弱,而 MTG 染色持续存在,表明线粒体在焦亡过程中去极化。图 1. 细胞焦亡对线粒体损伤的影响[1]。 (A) 用 MitoTracker 深红 (MTDR)、MitoTracker 绿 (MTG) 和 PI 染色的 LPS + Nigericin 处理的 THP1 的共聚焦荧光活细胞图像。白色箭头表示焦亡细胞。(B) 黑色箭头表示正常线粒体;黄色箭头,线粒体嵴缺失或膜损伤;红色箭头,自噬体内受损的线粒体。
同时,在添加 Nigericin 引发焦亡前后,用透射电子显微镜观察 LPS 原生的 THP-1 细胞,可以看到线粒体形态发生了变化 (图 1C-D):
- 添加前:线粒体呈典型的卵形,由双膜囊泡和内部嵴状结构组成,但在添加后,线粒体收缩并圆润起来,平均长度减少约 2 倍 。
- 添加后 15 min 内,观察到含有损伤线粒体的自噬体。1 h 后,约 60% 的线粒体具有旋转或消失的嵴状结构和/或破裂的内线粒体膜 (IMM) 和外线粒体膜 (OMM),线粒体数量减少约 2 倍。
在引发焦亡后的早期阶段,线粒体受到了严重损害。此外,活化的 GSDMD-NT 会转位至线粒体并在其膜上成孔,导至线粒体膜通透。
▐ 细胞先发生焦亡?还是线粒体损伤 ?
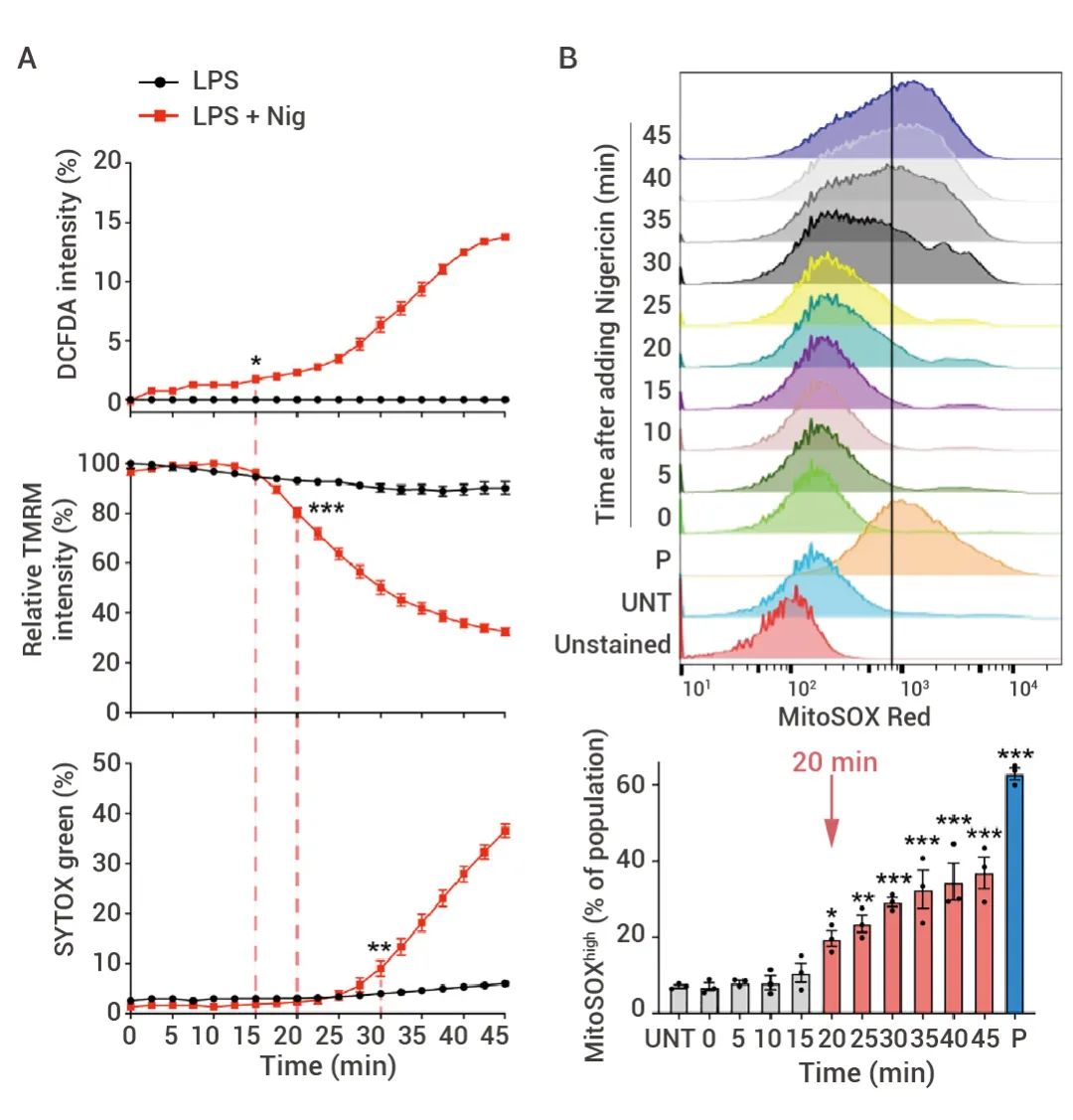
研究人员评估了 LPS + Nigericin 处理过的 THP-1 细胞中的线粒体膜电位 (TMRM 强度)、细胞 ROS 产生 (DCFDA 强度) 和质膜通透性 (SYTOX Green摄取) (图 2A)。DCFDA 增加而 TMRM 降低发生在细胞摄取 SYTOX Green 之前,证实了线粒体功能不全在细胞死亡之前发生。流式细胞术对 MitoSOX (线粒体 ROS)、DiIC1(5) (线粒体膜电位) 和 SYTOX Green 摄取进行的检验也证实了这一点 (图 2B)。在 SYTOX Green 摄取之前约 10 min 就能检测到 MitoSOX 和 DiIC1(5) 的显著变化。
图 2. 线粒体损伤发生在质膜损伤前,并增强焦亡[1]。
(A) 利用酶标仪对 LPS 或 LPS+Nigericin (Ng) 处理的 THP-1 中的 DCF 和 TRM 强度以及 SYTOX Green 摄取进行动力学分析。(B) LPS+Nigericin (Ng) 处理的 THP1 处理后用 MitoSOX、DiIC1(5) 或 SYTOX Green 染色的的流式细胞术直方图(上)和多个样品的定量(下)分析结果。P,阳性对照(抗霉素 A处理检测 MitoSOX,CCCP 处理检测 DiIC1(5),Triton X-100 处理检测 SYTOX Green);UNT,未经治疗。
同时,使用溴化乙锭 (EB) 处理 iBMDMs (永生化小鼠骨髓来源巨噬细胞) 以生成缺少线粒体的 ρ° 细胞。经检测,发现 ρ° 细胞对于经典及非经典的焦亡途径炎症小体诱导的焦亡具有抗性,线粒体缺陷会显著降低细胞焦亡诱导,表明线粒体在焦亡中发挥关键作用。此外,胞质 ROS 可放大经典和非经典炎症小体介导的细胞焦亡。
 ▐ GSDMD-NT 在质膜破裂前转移并损伤线粒体 ▐ GSDMD-NT 在质膜破裂前转移并损伤线粒体
为了确定在活细胞中 GSDMD-NT 是否结合并介导线粒体功能障碍,我们对表达多西环素 (DOX) 诱导的 I105N GSDMD-NT 的 iBMDMs 进行了成像,该 GSDMD-NT 在其 C 末端融合了蓝色荧光蛋白 (GSDMD-NT-BFP)。I105N 突变使焦亡速度减慢,从而使得更好地检测到死亡细胞。
I105N GSDMD-NT-BFP 的 iBMDMs:
iBMDMs, Immortalized bone marrow derived macrophages,永生化骨髓源性巨噬细胞。作者使用了来自表达 Cas9 的小鼠的永生化骨髓源性巨噬细胞 (iBMDM),即 Cas9 KI iBMDM。
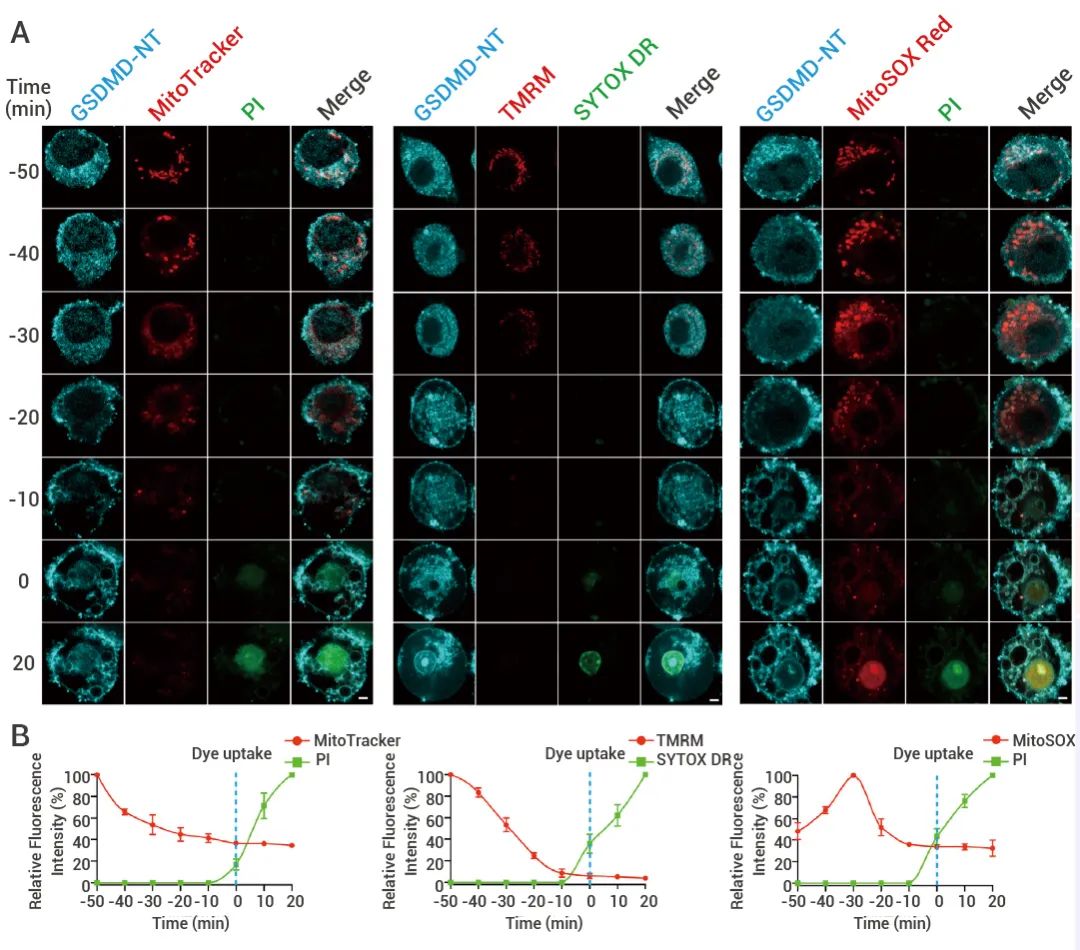
作者在 Cas9 KI iBMDM 中稳定表达了 Tet3G 反式激活子,以及在 Cas9 KI Tet3G 稳定细胞中产生多西环素 (Dox) 诱导型 GSDMD 变体,这使得编码 NT-GSDMD 和全长 GSDMD (FL-GSDMD) 的转基因能够通过Dox 诱导表达。其中包含 C 端 BFP 标签。利用了包含 I105N 突变的 NT-GSDMD 变体 (已被证明可以防止巨噬细胞焦亡),因为这种突变体比其野生型 (WT) 对应物更容易在细胞内检测到[7][8]。研究表明早期的 GSDMD-NT定位到线粒体 (图 3 A-E)。在 DOX 诱导后 4-8 h,可以检测到 GSDMD-NT-BFP。通过活细胞共聚焦成像观察了添加 DOX 后 6 小时开始的 GSDMD-NT 定位、质膜通透性 (PI 或 SYTOX 摄取) 和线粒体损伤 (MitoTracker、TMRM 和 MitoSOX) 的动态 (图 3A,3B)。
图 3. GSDMD-NT 在质膜前易位并损伤线粒体[1]。 (A 和 B) 通过DOX 诱导稳定表达 GSDMD-NT(I105N)-BFP 的 iBMDM,利用MitoTracker Deep Red 和 PI(左)、TMRM 和 SYTOX Deep Red (SYTOX DR)(中)或 MitoSOX Red 和 PI(右) 进行活细胞成像用评估线粒体损伤和细胞死亡,在添加 DOX 后 6 小时开始成像。时间 0 表示首次检测到细胞中的 PI 或 SYTOX(蓝色虚线)。
通过定义每个细胞在首次检测到染料摄取 (质膜损伤) 时的时间为 0 来同步焦亡变化:在 DOX 后,最终经 PI 或 SYTOX 摄取评估死亡的 GSDMD-NT-BFP 表达细胞中,MitoTracker 和 TMRM 强度都降低了(图 3A 和 3B),而在表达 FL-GSDMD-BFP 的细胞中,线粒体染料强度和 PI 摄取保持不变 (图 S2C-2F)。GSDMD-NT-BFP 至少在质膜通透性增加前 50 分钟就与线粒体染料定位在一起 (图3A 和 3B),确认了早期的 GSDMD-NT 定位到线粒体。当然,作者也通过进一步的成像实验证实 GSDMD-NT 与线粒体的共定位 (此处不再详细展开)。
插一句:换个思路想,这就好比谈恋爱, A 和 B 分手了,B 立刻就和 C 在一起了,说明啥?B 和 C 肯定早就在一起了嘛!哈哈哈哈仅供理解,不可细究喔~
此外,MitoTracker 和 TMRM 强度逐渐下降,而 MitoSOX 信号在 PI 或 SYTOX 摄取之前增加。所有线粒体标记最终消失,暗示线粒体溶解。在焦亡晚期,质膜已经被渗透后,从受损线粒体释放出 MitoSOX Red。也就是说,细胞焦亡早期,GSDMD-NT 会首先转位至线粒体,且在细胞膜成孔之前,造成线粒体损伤。
▐ GSDMD-NT 直接使 OMM 和 IMM 通透
为了确定 GSDMD-NT 是否自身就能结合并损伤线粒体膜,作者从 HCT116 (A-C)或 iBMDMs (D-E) 中分离出线粒体,在存在或不存在 z-VAD-FMK (泛 Caspase 抑制剂) 或 Disulfiram (DSF, GSDMD 孔形成的有效抑制剂) 的情况下,用 GSDMD 和/或 Caspase-11 或 t-Bid 处理。GSDMD 和 Caspase-11 处理可产生活性的 GSDMD-NT,并通过免疫印迹检测释放的线粒体蛋白以评估线粒体通透性 (图 4A 和 4B)。
图 4. GSDMD-NT 可通透并损伤分离的线粒体[1]。 (A、B 和 D) 处理后通过免疫印迹检测上清液或线粒体的蛋白 (A 和 D) 以及多个实验的光密度测定 (B)。(C 和 E) 通过 qPCR 检测上清液中的 mtDNA。
研究表明,GSDMD-NT 直接使 OMM 和 IMM 通透,而不需要其他因素。- 添加 Caspase-11 和 GSDMD 后 5 分钟内,可溶性线粒体基质蛋白 ACO2 和线粒体膜间隙蛋白 (IMS 蛋白:Cytochrome C 和 HtrA2) 从线粒体中释放出来,而与膜结合的 COX IV 则没有。泛 Caspase 抑制剂 z-VAD-FMK 抑制了释放。
- 在添加 t-Bid 后,触发凋亡中的 MOMP (线粒体外膜通透化) 而不破坏 IMM (线粒体内膜),释放出了 Cytochrome C 和 HtrA2,但没有释放 ACO2 (图 4A-B)。
- 当线粒体与 GSDMD 和 Caspase-11 一起孵育时,也释放出 mtDNA,但是单独添加它们或者添加 t-Bid 时则没有 (图 4C)。
- 同时,处理分离的线粒体的 GSDMD 和 Caspase-11 也显著增加了 ROS,并通过 MitoSOX Red 和 DiIC1(5) 染色分别降低了膜电位 (图中未显示)。
此外,Disulfiram 抑制 GSDMD 孔洞形成,但不干扰 GSDMD 切割或 GSDMD-NT 与膜的结合。在添加 GSDMD 和 Caspase-11 之前预先用 DSF 孵育分离的线粒体 (图 4D-E)。DSF 阻止了 Cytochrome C 和 mtDNA 的线粒体释放,证实了 GSDMD-NT 孔洞使线粒体通透。
▐ OMM 心磷脂是 GSDMD 介导的线粒体损伤所必须的
心磷脂是由 CRLS1 合成并由 PLSCR3 外化到外线粒体膜 (OMM) 上 (图 5 A),为研究 GSDMD-NT 介导的线粒体损伤是否需要 OMM 心磷脂,作者对 iBMDMs 中对 CRLS1 和 PLSCR3 进行了基因敲除 (Cris1-/-和 Plscr3-/- iBMDMs),并用 LPS 和 Nigericin 进行处理。
图 5. OMM心磷脂是GSDMD介导的线粒体损伤所必需[1]。 (A) 心磷脂合成和易位示意图。PG,磷脂酰甘油;CDP-DAG,二磷酸胞苷-二酰基甘油。(B) LPS或 LPS+Nigericin 处理的表达 mNG-GSDMD 的 WT、Plscr3-/-或 Crlsr1-/- iBMDM 的共聚焦活细胞成像,在添加 Nigericin 30 分钟后进行 MitoTracker 深红染色。白色箭头表示 GSDMD 的线粒体募集。(C) 未处理或Nigerici n处理 30 分钟后,LPS 预处理的 iBMDM 的全细胞裂解物 (WCL)、胞质 (细胞) 和线粒体 (mito)的免疫印迹。 (D) 通过 qPCR 检测胞质 mtDNA。
研究发现, OMM 心磷脂介导了 GSDMD 依赖性的线粒体损伤。- 在 LPS+Nigericin 处理的 Cris1-/- 和 Plscr3-/- iBMDMs 中,GSDMD-NT 没有被招募到线粒体 (图 5B),而线粒体完整性和膜电位、线粒体数量和平均长度以及损害线粒体的百分比几乎恢复到未处理的 WT iBMDMs 的状态 (图中未显示)。
- 此外,线粒体 ROS (图中未显示) 和 Cytochrome C 和 mtDNA 的胞质释放 (图5C-D) 也被阻止。一致地,用 GSDMD 和 Caspase-11 处理的 Crls1-/- 和Plscr3-/- iBMDMs 的分离线粒体没有释放 Cytochrome C、HtrA2 或 mtDNA (图中未显示)。
同时,GSDMD 介导的线粒体损伤诱导 3’端- 5’端的核酸外切酶 PNPT1 释放入胞质,引发广泛的 mRNA 降解以加重细胞焦亡发生、放大下游炎性反应。也正是如此,提供了一个强大的正反馈机制来放大细胞死亡,被称为“不归路”。此外,该研究还揭示了线粒体损伤增强细胞焦亡诱导的机体抗肿瘤免疫应答。
 LPS + Nigericin
诱导 THP-1 细胞焦亡。 | MitoTracker Deep Red / MitoTracker Green
线粒体红色/绿色荧光探针。 | TMRM
膜渗透性的阳离子荧光探针,检测线粒体膜电位。 | DCFDA
具有细胞渗透性,可以检测细胞内的 ROS。 | MitoTEMPO
是一种靶向线粒体的超氧化物歧化酶模拟物,有着清除超氧化物和烷基自由基的能力。 | MVRGreen
一种可渗透的 DNA 染料。
细胞焦亡化合物库
收录了 1186 种细胞焦亡相关产品,可以用于细胞焦亡信号通路及相关疾病的研究。 |
MCE的所有产品仅用作科学研究或药证申报,我们不为任何个人用途提供产品和服务。
参考文献:
[1] Rui Miao, et al. Gasdermin D permeabilization of mitochondrial inner and outer membranes accelerates and enhances pyroptosis. Immunity. 2023 Nov 14;56(11):2523-2541.e8.
[2] Dominic P Del Re, et al. Fundamental Mechanisms of Regulated Cell Death and Implications for Heart Disease. Physiol Rev. 2019 Oct 1;99(4):1765-1817.
[3] Tessa Bergsbaken, et al. Pyroptosis: host cell death and inflammation. Nat Rev Microbiol. 2009 Feb;7(2):99-109.
[4] Xing Liu, et al. Inflammasome-activated gasdermin D causes pyroptosis by forming membrane pores. Nature. 2016 Jul 7;535(7610):153-8.
[5] Wei X,et al. Role of pyroptosis in inflammation and cancer. Cell Mol Immunol. 2022 Sep;19(9):971-992.
[6] Jibo Han, et al. GSDMD (Gasdermin D) Mediates Pathological Cardiac Hypertrophy and Generates a Feed-Forward Amplification Cascade via Mitochondria-STING (Stimulator of Interferon Genes) Axis. Hypertension. 2022 Nov;79(11):2505-2518.
[7] Aglietti RA,et al. GsdmD p30 elicited by caspase-11 during pyroptosis forms pores in membranes. Proc Natl Acad Sci U S A. 2016 Jul 12;113(28):7858-63.
[8] Evavold CL, et al. Control of gasdermin D oligomerization and pyroptosis by the Ragulator-Rag-mTORC1 pathway. Cell. 2021 Aug 19;184(17):4495-4511.e19.
|

 /3
/3 

 浙公网安备33010802005999号
浙公网安备33010802005999号 






 2026庆【网站十三周
2026庆【网站十三周 2025庆【网站十二周
2025庆【网站十二周 2024庆中秋、迎国庆
2024庆中秋、迎国庆 2024庆【网站十一周
2024庆【网站十一周 2023庆【网站十周年
2023庆【网站十周年 2022庆【网站九周年
2022庆【网站九周年

 发表于 2024-3-25 16:17
发表于 2024-3-25 16:17